CATEGORIES:
BiologyChemistryConstructionCultureEcologyEconomyElectronicsFinanceGeographyHistoryInformaticsLawMathematicsMechanicsMedicineOtherPedagogyPhilosophyPhysicsPolicyPsychologySociologySportTourism
Relation to evolution
Ecology and evolution are considered sister disciplines of the life sciences. Natural selection, life history, development, adaptation, populations, and inheritance are examples of concepts that thread equally into ecological and evolutionary theory. Morphological, behavioral and/or genetic traits, for example, can be mapped onto evolutionary trees to study the historical development of a species in relation to their functions and roles in different ecological circumstances. In this framework, the analytical tools of ecologists and evolutionists overlap as they organize, classify and investigate life through common systematic principals, such as phylogenetics or the Linnaean system of taxonomy.[116] The two disciplines often appear together, such as in the title of the journal Trends in Ecology and Evolution.[117] There is no sharp boundary separating ecology from evolution and they differ more in their areas of applied focus. Both disciplines discover and explain emergent and unique properties and processes operating across different spatial or temporal scales of organization.[50][118][119] While the boundary between ecology and evolution is not always clear, it is understood that ecologists study the abiotic and biotic factors that influence the evolutionary process.[120][121]
Behavioral ecology
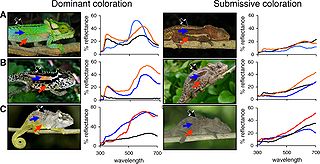

Social display and color variation in differently adapted species of chameleons (Bradypodion spp.). Chameleons change their skin color to match their background as a behavioral defense mechanism and also use color to communicate with other members of their species, such as dominant (left) versus submissive (right) patterns shown in the three species (A-C) above.[122]
All organisms are motile to some extent. Even plants express complex behavior, including memory and communication.[123] Behavioral ecology is the study of an organism's behavior in its environment and its ecological and evolutionary implications. Ethology is the study of observable movement or behavior in animals. This could include investigations of motile sperm of plants, mobile phytoplankton, zooplankton swimming toward the female egg, the cultivation of fungi by weevils, the mating dance of a salamander, or social gatherings of amoeba.[124][125][126][127][128]
Adaptation is the central unifying concept in behavioral ecology.[129] Behaviors can be recorded as traits and inherited in much the same way that eye and hair color can. Behaviors evolve and become adapted to the ecosystem because they are subject to the forces of natural selection.[15] Hence, behaviors can be adaptive, meaning that they evolve functional utilities that increases reproductive success for the individuals that inherit such traits.[130] This is also the technical definition for fitness in biology, which is a measure of reproductive success over successive generations.[15]
Predator-prey interactions are an introductory concept into food-web studies as well as behavioral ecology.[131] Prey species can exhibit different kinds of behavioral adaptations to predators, such as avoid, flee or defend. Many prey species are faced with multiple predators that differ in the degree of danger posed. To be adapted to their environment and face predatory threats, organisms must balance their energy budgets as they invest in different aspects of their life history, such as growth, feeding, mating, socializing, or modifying their habitat. Hypotheses posited in behavioral ecology are generally based on adaptive principals of conservation, optimization or efficiency.[34][120][132] For example,
"The threat-sensitive predator avoidance hypothesis predicts that prey should assess the degree of threat posed by different predators and match their behavior according to current levels of risk."[133]
"The optimal flight initiation distance occurs where expected postencounter fitness is maximized, which depends on the prey's initial fitness, benefits obtainable by not fleeing, energetic escape costs, and expected fitness loss due to predation risk."[134]

Symbiosis: Leafhoppers (Eurymela fenestrata) are protected by ants (Iridomyrmex purpureus) in a symbiotic relationship. The ants protect the leafhoppers from predators and in return the leafhoppers feeding on plants exude honeydew from their anus that provides energy and nutrients to tending ants.[135]
Elaborate sexual displays and posturing are encountered in the behavioral ecology of animals. The birds of paradise, for example, display elaborate ornaments and song during courtship. These displays serve a dual purpose of signaling healthy or well-adapted individuals and desirable genes. The elaborate displays are driven by sexual selection as an advertisement of quality of traits among male suitors.[136]
Social ecology
Social ecological behaviors are notable in the social insects, slime moulds, social spiders, human society, and naked mole rats where eusocialism has evolved. Social behaviors include reciprocally beneficial behaviors among kin and nest mates.[15][126][137] Social behaviors evolve from kin and group selection. Kin selection explains altruism through genetic relationships, whereby an altruistic behavior leading to death is rewarded by the survival of genetic copies distributed among surviving relatives. The social insects, including ants, bees and wasps are most famously studied for this type of relationship because the male drones are clones that share the same genetic make-up as every other male in the colony.[15] In contrast, group selectionists find examples of altruism among non-genetic relatives and explain this through selection acting on the group, whereby it becomes selectively advantageous for groups if their members express altruistic behaviors to one another. Groups that are predominantly altruists beat groups that are predominantly selfish.[15][138]
Coevolution
Ecological interactions can be divided into host and associate relationships. A host is any entity that harbors another that is called the associate.[139] Host and associate relationships among species that are mutually or reciprocally beneficial are called mutualisms. If the host and associate are physically connected, the relationship is called symbiosis. Approximately 60% of all plants, for example, have a symbiotic relationship with arbuscular mycorrhizal fungi. Symbiotic plants and fungi exchange carbohydrates for mineral nutrients.[140] Symbiosis differs from indirect mutualisms where the organisms live apart. For example, tropical rainforests regulate the Earth's atmosphere. Trees living in the equatorial regions of the planet supply oxygen into the atmosphere that sustains species living in distant polar regions of the planet. This relationship is called commensalism because many other host species receive the benefits of clean air at no cost or harm to the associate tree species supplying the oxygen.[141] The host and associate relationship is called parasitism if one species benefits while the other suffers. Competition among species or among members of the same species is defined as reciprocal antagonism, such as grasses competing for growth space.[142]

Parasites: A harvestman arachnid is parasitized by mites. This is parasitism because the harvestman is being consumed as its juices are slowly sucked out while the mites gain all the benefits traveling on and feeding off of their host.
Popular ecological study systems for mutualism include, fungus-growing ants employing agricultural symbiosis, bacteria living in the guts of insects and other organisms, the fig wasp and yucca moth pollination complex, lichens with fungi and photosynthetic algae, and corals with photosynthetic algae.[143][144] Nevertheless, many organisms exploit host rewards without reciprocating and thus have been branded with a myriad of not-very-flattering names such as 'cheaters', 'exploiters', 'robbers', and 'thieves'. Although cheaters impose several host cots (e.g., via damage to their reproductive organs or propagules, denying the services of a beneficial partner), their net effect on host fitness is not necessarily negative and, thus, becomes difficult to forecast.[145][146]
Biogeography
The word biogeography is an amalgamation of biology and geography. Biogeography is the comparative study of the geographic distribution of organisms and the corresponding evolution of their traits in space and time.[147] The Journal of Biogeography was established in 1974.[148] Biogeography and ecology share many of their disciplinary roots. For example, the theory of island biogeography, published by the mathematician Robert MacArthur and ecologist Edward O. Wilson in 1967[149] is considered one of the fundamentals of ecological theory.[150]
Biogeography has a long history in the natural sciences where questions arise concerning the spatial distribution of plants and animals. Ecology and evolution provide the explanatory context for biogeographical studies.[147] Biogeographical patterns result from ecological processes that influence range distributions, such as migration and dispersal.[150] and from historical processes that split populations or species into different areas.[151] The biogeographic processes that result in the natural splitting of species explains much of the modern distribution of the Earth's biota. The splitting of lineages in a species is called vicariance biogeography and it is a sub-discipline of biogeography.[151][152][153] There are also practical applications in the field of biogeography concerning ecological systems and processes. For example, the range and distribution of biodiversity and invasive species responding to climate change is a serious concern and active area of research in context of global warming.[20][154]
r/K-Selection theory
A population ecology concept (introduced in MacArthur and Wilson's (1967) book, The Theory of Island Biogeography) is r/K selection theory, one of the first predictive models in ecology used to explain life-history evolution. The premise behind the r/K selection model is that natural selection pressures change according to population density. For example, when an island is first colonized, density of individuals is low. The initial increase in population size is not limited by competition, leaving an abundance of available resources for rapid population growth. These early phases of population growth experience density-independent forces of natural selection, which is called r-selection. As the population becomes more crowded, it approaches the island's carrying capacity, thus forcing individuals to compete more heavily for fewer available resources. Under crowded conditions the population experiences density-dependent forces of natural selection, called K-selection.[155]
In the r/K-selection model, the first variable r is the intrinsic rate of natural increase in population size and the second variable K is the carrying capacity of a population.[34] Different species evolve different life-history strategies spanning a continuum between these two selective forces. An r-selected species is one that has high birth rates, low levels of parental investment, and high rates of mortality before individuals reach maturity. Evolution favors high rates of fecundity in r-selected species. Many kinds of insects and invasive species exhibit r-selected characteristics. In contrast, a K-selected species has low rates of fecundity, high levels of parental investment in the young, and low rates of mortality as individuals mature. Humans and elephants are examples of species exhibiting K-selected characteristics, including longevity and efficiency in the conversion of more resources into fewer offspring.[149][156]
Molecular ecology
The important relationship between ecology and genetic inheritance predates modern techniques for molecular analysis. Molecular ecological research became more feasible with the development of rapid and accessible genetic technologies, such as the polymerase chain reaction (PCR). The rise of molecular technologies and influx of research questions into this new ecological field resulted in the publication Molecular Ecology in 1992.[157] Molecular ecology uses various analytical techniques to study genes in an evolutionary and ecological context. In 1994, John Avise also played a leading role in this area of science with the publication of his book, Molecular Markers, Natural History and Evolution.[158] Newer technologies opened a wave of genetic analysis into organisms once difficult to study from an ecological or evolutionary standpoint, such as bacteria, fungi and nematodes. Molecular ecology engendered a new research paradigm for investigating ecological questions considered otherwise intractable. Molecular investigations revealed previously obscured details in the tiny intricacies of nature and improved resolution into probing questions about behavioral and biogeographical ecology.[158] For example, molecular ecology revealed promiscuous sexual behavior and multiple male partners in tree swallows previously thought to be socially monogamous.[159] In a biogeographical context, the marriage between genetics, ecology and evolution resulted in a new sub-discipline called phylogeography.[160]
Human ecology
Human ecology is the interdisciplinary investigation into the ecology of our species. "Human ecology may be defined: (1) from a bio-ecological standpoint as the study of man as the ecological dominant in plant and animal communities and systems; (2) from a bio-ecological standpoint as simply another animal affecting and being affected by his physical environment; and (3) as a human being, somehow different from animal life in general, interacting with physical and modified environments in a distinctive and creative way. A truly interdisciplinary human ecology will most likely address itself to all three."[161] The term human ecology was formally introduced in 1921, but many sociologists, geographers, psychologists, and other disciplines were interested in human relations to natural systems centuries prior, especially in the late 19th century.[161][162] Some authors have identified a new unifying science in coupled human and natural systems that builds upon, but moves beyond the field human ecology.[163] Ecology is as much a biological science as it is a human science.[6] "Perhaps the most important implication involves our view of human society. Homo sapiens is not an external disturbance, it is a keystone species within the system. In the long term, it may not be the magnitude of extracted goods and services that will determine sustainability. It may well be our disruption of ecological recovery and stability mechanisms that determines system collapse."[72]:3282
Date: 2016-01-03; view: 3683
| <== previous page | | | next page ==> |
| Ecological complexity | | | Metabolism and the early atmosphere |