CATEGORIES:
BiologyChemistryConstructionCultureEcologyEconomyElectronicsFinanceGeographyHistoryInformaticsLawMathematicsMechanicsMedicineOtherPedagogyPhilosophyPhysicsPolicyPsychologySociologySportTourism
Geological Record of the Cambrian Explosion
The geological record provides an important clue to the origin of biomineralization in the animal phyla, as has been noted by many previous authors [5-12]. Prior to the Cambrian (~ 544 Myr ago), almost the only evidence for animal life was the softbodied Ediacaran fauna (Fig. 10.1). Together with related bed-parallel tracks
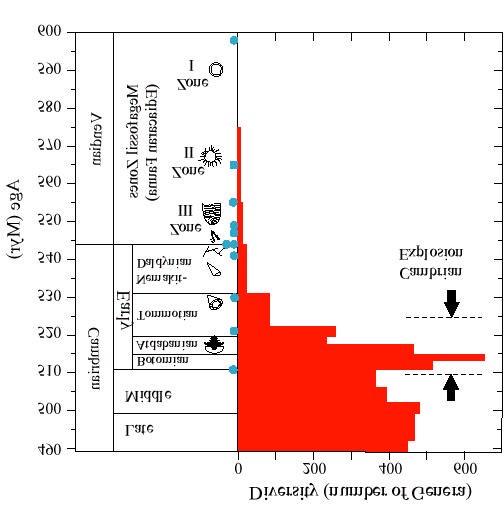
Figure 10.1. Radiometric age constraints and generic diversity for Vendian and Cambrian time. Position of the actual U/Pb constraints are indicated by solid blue dots. Data for the generic diversity have been compiled from Narbonne et al. [27] and Sepkoski [28], and are plotted as the number of taxa reported in each reference time interval. The black arrows indicate the approximate time of the Cambrian Explosion / true polar wander event [31]. The original figure was adapted from that of Grotzinger et al. [25].
and trails of this interval, these extraordinarily rare fossils are typically preserved as casts/molds at bed interfaces, and display no evidence of biomineralization. Molecular clock studies of protein divergence times for the major animal phyla have indicated consistently that major phyla separated up to several hundred mil-lion years prior to their first appearance in the fossil record [13-18]. The first clear evidence of matrix-mediated Ca-biomineralization is the latest Precambrian invertebrate Cloudina [19] (Fig. 10.2). Extensive work on the numerical calibration of the geological time scale for this interval illustrates that Cloudina first appeared about 550 Myr ago, with other mineralized forms appearing during the next 40 Myr [20-26]. However, the largest burst of new biomineralization activity is clearly in the Tommotian/Atdabanian interval, where the diversity of fossil organisms (mostly new biomineralizing groups) increases nearly exponentially over a ~10 Myr interval.
The trigger for this Cambrian Explosion has been a subject of extensive debate in the geological literature. It seems to have been a time of general climatic insta-bility, as reflected by intense oscillations in the stable isotope record of carbon [17, 29, 30], probably driven by tectonic events of global magnitude [31]. Thus, it was a good time for new evolutionary innovations, because the climatic instability pro-vided opportunities for novel forms to become fixed within small populations. But the mineralization drive itself may have been triggered by a separate development: the evolution of an animal predator as suggested by Stanley [5]. Although there is no evidence for animal predation among Ediacaran paleocommunities, some of the earliest trace fossils associated with the Ediacarans appear to be scratch marks made by the hardened radula of an unidentified mollusc [32]. Similarly, the earliest skeletonized fossils (Cloudina) contain clear evidence of predatorial borings – borings likely to have been made by the rasping of a mineralized feeding apparatus [12]. It is therefore likely that the adaptive advantages conferred by skeletal and tooth biomineralization were an important factor influencing the intensity of the Cambrian explosion, and they may have been amplified by the co-evolution of predator/prey systems.
The mere presence of a good evolutionary driving force is not enough to com-pletely reinvent a complex biochemical system multiple times within a geologically short interval. Lowenstam and Margulis [6] noted that the vast majority of new biomineral products observed in the Early Cambrian were based on calcium - either some form of CaCO3, or Ca-phosphate minerals as shown on Fig. 10.2. Noting that precise control of intracellular calcium is necessary for the formation of the microtubules needed by all eukaryotic cells, they suggested that these calcium regulation and transport systems provided the evolutionary prerequisites for their eventual use in biomineralization (in effect, exaptation, although the word had not been coined in 1980). Calcium, carbonate, and phosphate ions are abundant in the world oceans, and hence would be favored for use in skeletons over much rarer materials like Fe, Sr, Mn, etc.
However, gathering the components is only a small part of the biologically controlled mineralization process. They need to be brought together in a confined volume, in a controlled fashion, and induced to crystallize. The growing crystallites need to be properly tended, fed, and confined to the desired size, shape, and crys-tallographic
orientation. Hence, it seems that another biochemical/genetic system,
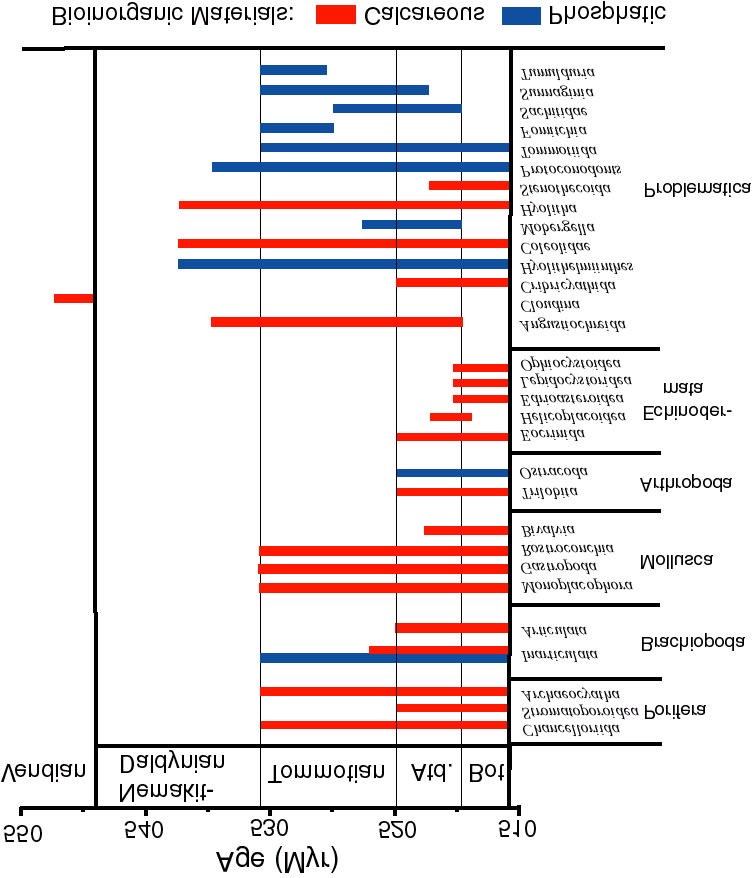
Figure 10.2. Stratigraphic ranges and first appearances of major fossil taxa that employ calcareous and phosphatic biomineralization. Not all families and problematic taxa are listed, nor are silica biomineralizing groups; for summaries of the stratigraphic range of these groups, see Bengtson [11]. Note that the mineralogy of Cloudina is poorly constrained; Grant [19] inferred a primary mineralogy of high-magnesian calcite based on preferential dolomitization of shell layers. Data from Lowenstam & Margolis [6] and Bengtson [11].
in addition to the ion transport system was involved in this exaptation which led to widespread biomineralization in the Early Cambrian. In all of these respects, the magnetite (Fe3O4) biomineralization system present in extant magnetotactic bacteria seems to fit as this missing link. As noted below, it is ancient, appears to be present in most of the animal phyla, and has all of the essential aspects of biologically controlled mineralization processes present in higher organisms. With apologies to physicists, it seems appropriate to dub this concept the “Grand Unified Theory of Biomineralization”. Unlike some physical ‘GUT’ theories, this one can be tested easily.
Date: 2015-02-03; view: 1743
| <== previous page | | | next page ==> |
| Introduction | | | Magnetite Biomineralization |