CATEGORIES:
BiologyChemistryConstructionCultureEcologyEconomyElectronicsFinanceGeographyHistoryInformaticsLawMathematicsMechanicsMedicineOtherPedagogyPhilosophyPhysicsPolicyPsychologySociologySportTourism
Cells: The Basic Units of Life
Just as atoms are the units of chemistry, cells are the building blocks of life. Three statements constitute the cell theory:
Cells are the fundamental units of life.
All organisms are composed of cells.
All cells come from preexisting cells.
Cells are composed of water molecules and the small and large molecules. Each cell contains at least 10,000 different types of molecules, most of them present in many copies. Cells use these molecules to transform matter and energy, to respond to their environment, and to reproduce themselves. The cell theory has three important implications. First, it means that studying cell biology is in some sense the same as studying life. The principles that underlie the functions of the single cell in a bacterium are similar to those governing the 60 trillion cells of your body. Second, it means that life is continuous. All those cells in your body came from a single cell, the fertilized egg, which came from the fusion of two cells, a sperm and an egg from your parents, whose cells came from their fertilized eggs, and so on. Finally, it means that the origin of life on Earth was marked by the origin of the first cells.
Cells may have come from stable bubbles
Isolation from the general environment can be achieved in the laboratory within aggregates produced from molecules made in prebiotic synthesis experiments. Called protobionts, these aggregates cannot reproduce, but they can maintain internal chemical environments that differ from their surroundings. Under the microscope, they look a lot like tiny cells. In the 1920s, Alexander Oparin mixed a large protein and a polysaccharide in solution. When he agitated this mixture, bubbles formed. He could also do this with other polymers. The interiors of these bubbles had much higher concentrations of the macromolecules than their surroundings. Moreover, they catalyzed chemical reactions, and they had some control over what left them and crossed the boundary into the environment. In other words, they were protobionts. Later, other researchers showed that if lipids are mixed in an aqueous environment, they spontaneously arrange themselves into droplets surrounded by a bilayer. Taken together with the prebiotic chemistry models and RNAworld hypothesis, these experiments suggest a bubble theory for the origin of cells.
Cell size is limited by the surface area-to-volume ratio
Most cells are tiny. The volume of cells ranges from 1 to 1,000 m3. The eggs of some birds are enormous exceptions, to be sure, and individual cells of several types of algae and bacteria are large enough to be viewed with the unaided eye. And although neurons (nerve cells) have a volume that is within the “normal” cell range, they often have fine projections that may extend for meters, carrying signals from one part of a large animal to another. But by and large, cells are minuscule. The reason for this relates to the change in the surface area-to-volume ratio (SA/V) of any object as it increases in size. As a cell increases in volume, its surface area also increases, but not to the same extent. This phenomenon has great biological significance for two reasons: The volume of a cell determines the amount of chemical activity it carries out per unit of time. The surface area of a cell determines the amount of substances the cell can take in from the outside environment and the amount of waste products it can release to the environment. As a living cell grows larger, its rate of waste production and its need for resources increase faster than its surface area. This explains why large organisms must consist of many small cells: cells are small in volume in order to maintain a large surface area-to-volume ratio. In a multicellular organism, the large surface area represented by the multitude of small cells that make up the organism enables it to carry out the multitude of functions required for survival. Special structures transport food, oxygen and waste materials to and from the small cells that are distant from the external surface of the organism.
Most cells are invisible to the human eye. The smallest object a person can typically discern is about 0.2 mm size. We refer to this measure as resolution, the distance apart two objects must be in order for them to be distinguished as separate; if they are closer together, they appear as a single blur. Many cells are much smaller than 0, 2 mm Microscopes are instruments used to improve resolution so that cells and their internal structures can be seen. There are two basic types of microscopes: light microscopes and electron microscopes. The light microscope (LM) uses glass lenses and visible light to form a magnified image of an object. It has a resolving power of about 0. 2 nano-m, which is 1,000 times that of the human eye. It allows visualization of cell sizes and shapes and some internal cell structures. The latter are hard to see under ordinary light, so cells are often killed and stained with various dyes to make certain structures stand out.
An electron microscope (EM) uses magnets to focus an electron beam, much as a light microscope uses glass lenses to focus a beam of light. Since we cannot see electrons, the electron microscope directs them at a fluorescent screen or photographic film to create a visible image. The resolving power of electron microscopes is about 0.5 nm, which is 400,000 times that of the human eye. This resolving power permits the details of many subcellular structures to be distinguished. Many techniques have been developed to enhance the views of cells we see under the light and electron microscopes.
Cells are surrounded by a plasma membrane
As we have noted, a plasma membrane separates each cell from its environment, creating a segregated (but not isolated) compartment. The plasma membrane is composed of a phospholipids bilayer, with the hydrophilic “heads” of the lipids facing the cell’s aqueous interior on one side of the membrane and the extracellular environment on the other. Proteins are embedded in the lipids. In many cases, these proteins protrude into the cytoplasm and into the extracellular environment. The plasma membrane allows the cell to maintain a more or less constant internal environment. A self-maintaining, constant internal environment is a key characteristic of life.
The plasma membrane acts as a selectively permeable barrier, preventing some substances from crossing while permitting other substances to enter and leave the cell. As the cell’s boundary with the outside environment, the plasma membrane is important in communicating with adjacent cells and receiving extracellular signals. The plasma membrane often has molecules protruding from it that are responsible for binding and adhering to adjacent cells.
Cells show two organizational patterns
Prokaryotic cell organization is characteristic of the domains Bacteria and Archaea. Organisms in these domains are called prokaryotes. Their cells do not have membrane-enclosed internal compartments. The first cells ever to form were undoubtedly similar in organization to modern prokaryotes.
Eukaryotic cell organization is found in the domain Eukarya, which includes the protists, plants, fungi, and animals.
The genetic material (DNA) of eukaryotic cells is contained in a special membrane-enclosed compartment called the nucleus. Eukaryotic cells also contain other membrane-enclosed compartments in which specific chemical reactions take place. Organisms with this type of cell organization are known as eukaryotes. Both prokaryotes and eukaryotes have prospered for many hundreds of millions of years of evolution and both are great success stories.
Prokaryotic Cells
Prokaryotes can live off more different and diverse energy sources than any other living creatures and they inhabit greater environmental extremes, such as very hot springs and very salty water. Prokaryotic cells are generally smaller than eukaryotic cells. Each prokaryote is a single cell, but many types of prokaryotes are usually seen in chains, small clusters, or even clusters containing hundreds of individuals. In this section, we will first consider the features that cells in the domains Bacteria and Archaea have in common. Then we will examine structural features that are found in some but not all prokaryotes.
Prokaryotic cells share certain features
All prokaryotic cells have the same basic structure:
The plasma membrane encloses the cell regulating the traffic of materials into and out of the cell and separating it from its environment.
A region called the nucleoid contains the hereditary material (DNA) of the cell.
The rest of the material enclosed in the plasma membrane is called the cytoplasm. The cytoplasm is composed of two parts: the liquid cytosol, and insoluble suspended particles including ribosomes.
The cytosol consists mostly of water that contains dissolved ions, small molecules, and soluble macromolecules such as proteins.
Ribosomes are granules about 25 nm in diameter that are sites of protein synthesis. The cytoplasm is not a static region. Rather, the substances in this aqueous environment are in constant motion. For example, a typical protein moves around the entire cell within a minute, and encounters many molecules along the way. Although structurally less complicated than eukaryotic cells, prokaryotic cells are functionally complex, carrying out thousands of biochemical transformations.
Some prokaryotic cells have specialized features
As they evolved, some prokaryotes developed specialized structures that gave a selective advantage to those cells that had them. These structures include a protective cell wall, an internal membrane for compartmentalization of chemical reactions and flagella for cell movement through the watery environment.
Cell walls
Most prokaryotes have a cell wall located outside the plasma membrane. The rigidity of the cell wall supports the cell and determines its shape. The cell walls of most bacteria, but not archaea, contain peptidoglycan, a polymer of amino sugars, cross-linked by covalent bonds to form a single giant molecule around the entire cell. In some bacteria, another layer—the outer membrane (a polysaccharide- rich phospholipid membrane)—encloses the peptidoglycan layer. Unlike the plasma membrane, this outer membrane is not a major permeability barrier, and some of its polysaccharides are disease-causing toxins. Enclosing the cell wall in some bacteria is a layer of slime composed mostly of polysaccharides and referred to as a capsule. The capsules of some bacteria may protect them from attack by white blood cells in the animals they infect. The capsule helps keep the cell from drying out, and sometimes it helps the bacterium attach to other cells. Many prokaryotes produce no capsule, and those that do have capsules can survive even if they lose them, so the capsule is not essential to cell life. As you will see later in this chapter, eukaryotic plant cells also have a cell wall, but it differs in composition and structure from the cell walls of prokaryotes.
Internal membranes
Some groups of bacteria—the cyanobacteria and some others—carry on photosynthesis. In these photosynthetic bacteria, the plasma membrane folds into the cytoplasm to form an internal membrane system that contains bacterial chlorophyll and other compounds needed for photosynthesis. The development of photosynthesis, probably by such internal membranes, was an important event in the early evolution of life on Earth. Other prokaryotes have internal membrane folds that remain attached to the plasma membrane. These mesosomes may function in cell division or in various energy-releasing reactions.
Flagella and pill
Some prokaryotes swim by using appendages called flagella. A single flagellum, made of a protein called flagellin, looks at times like a tiny corkscrew. It spins on its axis like a propeller, driving the cell along. Ring structures anchor the flagellum to the plasma membrane and, in some bacteria, to the outer membrane of the cell wall. The flagella cause the motion of the cell because if they are removed, the cell cannot move.
Cynoskeleton
Recent evidence suggests that some prokaryotes, especially rod-shaped bacteria, have an internal filamentous helical structure just below the plasma membrane. The proteins that make up this structure are similar in amino acid sequence to actin in eukaryotic cells, and since actin is part of the cytoskeleton in those cells (see below), it has been suggested that the helical filaments in prokaryotes play a role in cell shape.
Eukaryotic Cells
Animals, plants, fungi, and protists have cells that are usually larger and structurally more complex than those of the prokaryotes. Eukaryotic cells generally have dimensions ten times greater than those of prokaryotes; for example, the spherical yeast cell has a diameter of 8 m. Like prokaryotic cells, eukaryotic cells have a plasma membrane, cytoplasm, and ribosomes. But added on to this basic organization are compartments in the cytoplasm whose interiors are separated from the cytosol by a membrane.
Compartmentalization is the key to eukaryotic cell function
Some of the compartments in eukaryotic cells are like little factories that make specific products. Others are like power plants that take in energy in one form and convert it to a more useful form. These membranous compartments, as well as other structures (such as ribosomes) that lack membranes but possess distinctive shapes and functions, are called organelles. Each of these organelles has specific roles in its particular cell. These roles are defined by chemical reactions.
The nucleus contains most of the cell’s genetic material (DNA). The duplication of the genetic material and the first steps in decoding genetic information take place in the nucleus.
The mitochondrion is a power plant and industrial park, where energy stored in the bonds of carbohydrates is converted to a form more useful to the cell (ATP) and certain essential biochemical conversions of amino acids and fatty acids occur.
The endoplasmic reticulum and Golgi apparatus are compartments in which proteins are packaged and sent to appropriate locations in the cell.
Lysosomes and vacuoles are cellular digestive systems in which large molecules are hydrolyzed into usable monomers.
Chloroplasts perform photosynthesis. The membrane surrounding each organelle does two essential things: First, it keeps the organelle’s molecules away from other molecules in the cell with which they might react inappropriately. Second, it acts as a traffic regulator, letting important raw materials into the organelle and releasing its products to the cytoplasm. The evolution of compartmentalization was an important development in the ability of eukaryotic cells to specialize, forming the organs and tissues of a complex body.
Organelles can be studied by microscopy or isolated for chemical analysis
Cell organelles were first detected by light and electron microscopy. The use of stains targeted to specific macromolecules has allowed cell biologists to determine the chemical compositions of organelles. Besides microscopy, another way to look at cells is to take them apart. Cell fractionation begins with the destruction of the cell membrane. This allows the cytoplasmic components to flow out into a test tube. The various organelles can then be separated from one another on the basis of size or density. Biochemical analyses can then be done on the isolated organelles. Microscopy and cell fractionation have complemented each other, giving a complete picture of the structure and function of each organelle.
Organelles that Process Information
Living things depend on accurate, appropriate information— internal signals, environmental cues, and stored instructions— to respond appropriately to changing conditions and maintain a constant internal environment. In the cell, information is stored in the sequence of nucleotides in DNA molecules. Most of the DNA in eukaryotic cells resides in the nucleus. Information is translated from the language of DNA into the language of proteins at the ribosomes.
The nucleus contains most of the cell’s DNA
The single nucleus is usually the largest organelle in a cell. The nucleus of most animal cells is approximately 5 m in diameter—substantially larger than most entire prokaryotic cells. The nucleus is the site of DNA duplication. The nucleus is the site of genetic control of the cell’s activities. A region within the nucleus, the nucleolus, begins the assembly of ribosomes from specific proteins and RNA. The nucleus is surrounded by two membranes, which together form the nuclear envelope. The two membranes of the nuclear envelope are separated by 10–20 nm and are perforated by nuclear pores approximately 9 nm in diameter, which connect the interior of the nucleus with the cytoplasm. At these pores, the outer membrane of the nuclear envelope is continuous with the inner membrane. Each pore is surrounded by a pore complex made up of eight large protein granules arranged in an octagon where the inner and outer membranes merge.
RNA and proteins pass through these pores to enter or leave the nucleus. At certain sites, the outer membrane of the nuclear envelope folds outward into the cytoplasm and is continuous with the membrane of another organelle, the endoplasmic reticulum (discussed later in this chapter). Inside the nucleus, DNA combines with proteins to form a fibrous complex called chromatin. Chromatin consists of exceedingly long, thin, entangled threads. Prior to cell division, the chromatin aggregates to form discrete, readily visible structures called chromosomes. Surrounding the chromatin are water and dissolved substances collectively referred to as the nucleoplasm. Within the nucleoplasm, a network of apparently structural proteins called the nuclear matrix organizes the chromatin. At the periphery of the nucleus, the chromatin is attached to a protein meshwork, called the nuclear lamina, which is formed by the polymerization of proteins called lamins into filaments. The nuclear lamina maintains the shape of the nucleus by its attachment to both the chromatin and the nuclear envelope. During most of a cell’s life cycle, the nuclear envelope is astable structure. When the cell divides, however, the nuclearenvelope fragments into pieces of membrane with attached pore complexes. The envelope re-forms when distribution ofthe duplicated DNA to the daughter cells is completed.
Ribosomes are the sites of protein synthesis
In prokaryotic cells, ribosomes float freely in the cytoplasm.In eukaryotic cells they occur in two places: in the cytoplasm,where they may be free or attached to the surface of the endoplasmic reticulum (described in the next section); and inside the mitochondria and chloroplasts, where energy is processed. In each of these locations, the ribosomes are the sites where proteins are synthesized under the direction of nucleic acids. Although they seem small in comparison to the cell in which they are contained, ribosomes are huge machines made up of several dozen kinds of molecules. The ribosomes of prokaryotes and eukaryotes are similar in that both consist of two different-sized subunits. Eukaryotic ribosomes are somewhat larger but the structure of prokaryotic ribosomes is better understood. Chemically, ribosomes consist of a special type of RNA, called ribosomal RNA (rRNA), to which more than 50 different protein moleculesare noncovalently bound.
The Endomembrane System
Much of the volume of some eukaryotic cells is taken up by an extensive endomembrane system. This system includes two main components, the endoplasmic reticulum and the Golgi apparatus. Continuities between the nuclear envelope and the endomembrane system are visible under the electron microscope. Tiny, membrane-surrounded droplets called vesicles appear to shuttle between the various components of the endomembrane system. This system has various structures but all of them are essentially compartments, closed off by their membranes from the cytoplasm. In this section, we will examine the functional significance of these compartments and we will see how materials synthesized in one organelle, the endoplasmic reticulum, are transferred to another organelle, the Golgi apparatus, for further processing, storage or transport. We will also describe the role of the lysosome in cellular digestion.
The endoplasmic reticulum is a complex factory
Electron micrographs reveal a network of interconnected membranes branching throughout the cytoplasm of a eukaryotic cell, forming tubes and flattened sacs. These membranes are collectively called the endoplasmic reticulum, or ER. The interior compartment of the ER, referred to as the lumen, is separate and distinct from the surrounding cytoplasm. The ER can enclose up to 10 percent of the interior volume of the cell, and its foldings result in a surface area many times greater than that of the plasma membrane.
Parts of the ER are studded with ribosomes, which are temporarily attached to the outer faces of its flattened sacs. These regions are called rough endoplasmic reticulum, or RER.
As a compartment, it segregates certain newly synthesized proteins away from the cytoplasm and transports them to other locations in the cell. While inside the RER, proteins can be chemically modified so as to alter their function and eventual destination. The attached ribosomes are sites for the synthesis of proteins that function outside the cytosol—that is, proteins that are to be exported from the cell incorporated into membranes or moved into the organelles of the endomembrane system. These proteins enter the lumen of the ER as they are synthesized. Once in the lumen of the ER, these proteins undergo several changes, including the formation of disulfide bridges and folding into their tertiary structures.
Proteins gain carbohydrate groups in the RER, thus becoming glycoproteins. In the case of proteins directed to the lysosomes, the carbohydrate groups are part of an “addressing” system that ensures that the right proteins are directed to the organelle.
Smooth endoplasmic reticulum or SER is more tubular (less like flattened sacs) and lacks ribosomes.
Within the lumen of the SER, proteins that have been synthesized on the RER are chemically modified. In addition, the SER has three other important roles:
It is responsible for chemically modifying small molecules taken in by the cell. This is especially true for drugs and pesticides.
It is the site for the hydrolysis of glycogen in animal cells.
It is the site for the synthesis of lipids and steroids. Cells that synthesize a lot of protein for export are usually packed with endoplasmic reticulum. Examples include glandular cells that secrete digestive enzymes and plasma cells that secrete antibodies. In contrast, cells that carry out less protein synthesis (such as storage cells) contain less ER. Liver cells which modify molecules that enter the body from the digestive system, have abundant smooth ER.
The Golgi apparatus stores,modifies, and packages proteins
The exact appearance of the Golgi apparatus (named for its discoverer, Camillo Golgi) varies from species to species but it always consists of flattened membranous sacs called cisternae and small membrane-enclosed vesicles. The cisternae appear to be lying together like a stack of saucers. The entire apparatus is about 1 m long.
The Golgi apparatus has several roles:
It receives proteins from the ER and may further modify them.
It concentrates, packages, and sorts proteins before they are sent to their cellular or extracellular destinations.
It is where some polysaccharides for the plant cell wall are synthesized.
In the cells of plants, protists, fungi, and many invertebrate animals, the stacks of cisternae are individual units scattered throughout the cytoplasm. In vertebrate cells, a few such stacks usually form a larger, single, more complex Golgi apparatus. The Golgi apparatus appears to have three functionally distinct parts: a bottom, a middle, and a top. The bottom cisternae, constituting the cis region of the Golgi apparatus, lie nearest to the nucleus or a patch of RER. The top cisternae constituting the trans region, lie closest to the surface of the cell. The cisternae in the middle make up the medial region of the complex. These three parts of the Golgi apparatus contain different enzymes and perform different functions. The Golgi apparatus receives proteins from the ER, packages them, and sends them on their way. Since there is often no direct membrane continuity between ER and Golgi apparatus, how does a protein get from one organelle to the other? The protein could simply leave the ER, travel across the cytoplasm, and enter the Golgi apparatus. But that would expose the protein to interactions with other molecules in the cytoplasm. On the other hand, segregation from the cytoplasm could be maintained if a piece of the ER could “bud off,” forming a vesicle that contains the protein— and that is exactly what happens. The protein makes the passage from ER to Golgi apparatus safely enclosed in the vesicle. Once it arrives, the vesicle fuses with the membrane of the Golgi apparatus, releasing its cargo. Vesicles form from the rough ER, move through the cytoplasm and fuse with the cis region of the Golgi apparatus, releasing their contents into the lumen. If living cells are stained specifically for ER and Golgi apparatus, the Golgi apparatus can be seen moving rapidly along the ER, possibly picking up vesicles as they go. Other small vesicles may move between the cisternae, transporting proteins, and it appears that some proteins move from one cisterna to the next by tiny channels. Vesicles budding off from the trans region carry their contents away from the complex.
Lysosomes contain digestive enzymes
Originating in part from the Golgi apparatus are organelles called lysosomes. They contain digestive enzymes, and they are the sites where macromolecules— proteins, polysaccharides, nucleic acids, and lipids—are hydrolyzed into their monomers. Lysosomes are about 1 m in diameter, are surrounded by a single membrane and have a densely staining, featureless interior. There may be dozens of lysosomes in a cell, depending on its needs. Lysosomes are sites for the breakdown of food and foreign objects taken up by the cell. These materials get into the cell by a process called phagocytosis (phago-, “eating”; cytosis, “cellular”), in which a pocket forms in the plasma membrane and eventually deepens and encloses material from outside the cell. This pocket becomes a small vesicle that breaks free of the plasma membrane to move into the cytoplasm as a phagosome containing food or other material. The phagosome fuses with a primary lysosome, forming a secondary lysosome where digestion occurs. The effect of this fusion is rather like releasing hungry foxes into a chicken coop: The enzymes in the secondary lysosome quickly hydrolyze the food particles. The products of digestion exit through the membrane of the lysosome, providing fuel molecules and raw materials for other cell processes. The “used” secondary lysosome, now containing undigested particles, then moves to the plasma membrane, fuses with it and releases the undigested contents to the environment. Lysosomes are also where the cell digests its own material in a process called autophagy. Autophagy is an ongoing process in which organelles such as mitochondria are engulfed by lysosomes and hydrolyzed to monomers, which pass out of the lysosome through its membrane into the cytoplasm for reuse. The importance of lysosome function is indicated by a group of human diseases called lysosomal storage diseases. If a cell lacks the ability to hydrolyze one or more macromolecules, these substances pile up in lysosomes, with harmful consequences. An example is Tay-Sachs disease, in which a lipid accumulates in the lysosomes of brain cells, resulting in death in early childhood. Plant cells do not appear to contain lysosomes, but the central vacuole of a plant cell (which we will describe below) may function in an equivalent capacity because it, like lysosomes, contains many digestive enzymes.
Organelles that Process Energy
A cell uses energy to synthesize cell-specific materials that it can use for activities such as growth, reproduction and movement. Energy is transformed from one form to another in mitochondria (found in all eukaryotic cells) and in chloroplasts (found in eukaryotic cells that harvest energy from sunlight). In contrast, energy transformations in prokaryotic cells are associated with enzymes attached to the inner surface of the plasma membrane or extensions of the plasma membrane that protrude into the cytoplasm.
Mitochondria are energy transformers
In eukaryotic cells, the breakdown of fuel molecules such as glucose begins in the cytosol. The molecules that result from this partial degradation enter the mitochondria (singular, mitochondrion), whose primary function is to convert the potential chemical energy of those fuel molecules into a form that the cell can use: the energy-rich molecule ATP (adenosine triphosphate). The production of ATP in the mitochondria using fuel molecules and molecular oxygen (O2) is called cellular respiration. Typical mitochondria are small—somewhat less than 1.5 µm in diameter and 2–8 µm in length—about the size of many bacteria. The number of mitochondria per cell ranges from one contorted giant in some unicellular protists to a few hundred thousand in large egg cells. An average human liver cell contains more than a thousand mitochondria. Cells that require the most chemical energy tend to have the most mitochondria per unit of volume. Mitochondria have two membranes. The outer membrane is smooth and protective, and it offers little resistance to the movement of substances into and out of the mitochondrion. Immediately inside the outer membrane is an inner membrane, which folds inward in many places, giving it a much greater surface area than that of the outer membrane. These folds tend to be quite regular, giving rise to shelflike structures called cristae.
The inner mitochondrial membrane contains many large protein molecules that participate in cellular respiration. The inner membrane exerts much more control over what enters and leaves the mitochondrion than does the outer membrane. The region enclosed by the inner membrane is referred to as the mitochondrial matrix. In addition to many proteins, the matrix contains some ribosomes and DNA that are used to make some of the proteins needed for cellular respiration.
Plastids photosynthesize or store materials
One class of organelles—the plastids—is produced only in plants and certain protists. There are several types of plastids, with different functions.
Chloroplasts
Chloroplasts contain the green pigment chlorophyll and are the sites of photosynthesis. In photosynthesis, light energy is converted into the chemical energy of bonds between atoms. The molecules formed in photosynthesis provide food for the photosynthetic organisms, as well as for other organisms that eat them. Directly or indirectly, photosynthesis is the energy source for most of the living world. Like a mitochondrion, a chloroplast is surrounded by two membranes. In addition, there is a series of internal membranes whose structure and arrangement vary from one group of photosynthetic organisms to another. Here we concentrate on the chloroplasts of the flowering plants. Even these chloroplasts show some variation but the pattern shown in Figure is typical. The internal membranes of chloroplasts look like stacks of flat, hollow pita bread. These stacks, called grana (singular, granum), consist of a series of flat, closely packed, circular compartments called thylakoids. In addition to phospholipids and proteins, the membranes of the thylakoids contain chlorophyll and other pigments that harvest light for photosynthesis. The thylakoids of one granum may be connected to those of other grana, making the interior of the chloroplast a highly developed network of membranes, much like the ER. The fluid in which the grana are suspended is the stroma. Like the mitochondrial matrix, the chloroplast stroma contains ribosomes and DNA, which are used to synthesize some, but not all, of the proteins that make up the chloroplast. Animal cells do not produce chloroplasts, but some do contain functional chloroplasts. These are either taken up as free chloroplasts derived from the partial digestion of green plants or contained within unicellular algae that live within the animal’s tissues. The green color of some corals and sea anemones results from the chloroplasts in algae that live within those animals. The animals derive some of their nutrition from the photosynthesis that their chloroplast-containing “guests” carry out. Such an intimate relationship between two different organisms is called symbiosis.
Other types of plastids also store pigments or polysaccharides:
Chromoplasts contain red, orange, and/or yellow pigments and give color to plant organs such as flowers. The chromoplasts have no known chemical function in the cell but the colors they give to some petals and fruits probably encourage animals to visit flowers and thus aid in pollination or to eat fruits and thus aid in seed dispersal. On the other hand, carrot roots gain no apparent advantage from being orange.
Leucoplasts are storage depots for starch and fats.
Endosymbiosis may explain the origin of mitochondria and chloroplasts
Although chloroplasts and mitochondria are about the size of prokaryotic cells and have the genetic material and protein synthesis machinery needed to make some of their own components, they are not independent of control by the nucleus. The vast majority of their proteins are encoded by nuclear DNA, made in the cytoplasm, and imported into the organelle.
Observations of these organelles have led to the proposal that they originated by endosymbiosis—that is, that they were once independent prokaryotic organisms. About 2 billion years ago, only prokaryotes inhabited Earth. Some of them absorbed their food directly from the environment. Others were photosynthetic. Still others fed on smaller prokaryotes by engulfing them. Suppose that a small, photosynthetic prokaryote was ingested by a larger one, but was not digested. Instead, it somehow survived, trapped within a vesicle in the cytoplasm of the larger cell. The smaller, ingested prokaryote divided at about the same rate as the larger one, so successive generations of the larger cell also contained the offspring of the smaller one. This phenomenon, called endosymbiosis (endo-, “within”; symbiosis, “living together”), exists today, as in the case of the algae that live within sea anemones (see Figure ). According to this scenario, endosymbiosis provided benefits for both partners: The larger cell obtained the photosynthetic products from the smaller cell, and the smaller cell was protected by the larger one. Over evolutionary time, the smaller cell gradually lost much of its DNA to the nucleus of the larger cell, resulting in the modern chloroplast. Much circumstantial evidence favors the endosymbiosistheory:
On an evolutionary time scale of millions of years, there is evidence for DNA moving between organelles in the cell.
There are many biochemical similarities between chloroplasts and modern photosynthetic bacteria.
DNA sequencing shows strong similarities between modern chloroplast DNA and that of a photosynthetic prokaryote.
The double membrane that encloses mitochondria and chloroplasts could have arisen through endosymbiosis.
The outer membrane may have come from the engulfing cell’s plasma membrane and the inner membrane from the engulfed cell’s plasma membrane. Similar evidence and arguments also support the proposition that mitochondria are the descendants of respiring prokaryotes prokaryotes engulfed by larger prokaryotes. The benefit of this endosymbiotic relationship might have been the capacity of the engulfed prokaryote to detoxify molecular oxygen (O2), which was increasing in Earth’s atmosphere as a result of photosynthesis.
Other Organelles
Eukaryotic cells have several other organelles that are surroundedby a single membrane.
Peroxisomes house specialized chemical reactions
Peroxisomes are organelles that collect the toxic peroxides (such as hydrogen peroxide, H2O2) that are the unavoidable by-products of chemical reactions. These peroxides can be safely broken down inside the peroxisomes without mixing with other parts of the cell. Peroxisomes are small organelles, about 0.2 to 1.7 µm in diameter. They have a single membrane and a granular interior containing specialized enzymes. Peroxisomes are found at one time or another in at least some of the cells of almost every eukaryotic species. Astructurally similar organelle, the glyoxysome, is found only in plants. Glyoxysomes, which are most prominent in young plants, are the sites where stored lipids are converted into carbohydrates for transport to growing cells.
Vacuoles are filled with water and soluble substances
Many eukaryotic cells, but particularly those of plants and protists, contain membrane-enclosed vacuoles filled with aqueous solutions containing many dissolved substances. Plant vacuoles have several functions:
Storage: Plant cells produce a number of toxic by-products and waste materials, many of which are simply stored within vacuoles. And since they are poisonous or distasteful, these stored materials deter some animals from eating the plants. Thus these stored wastes may contribute to plant survival.
Structure: In many plant cells, enormous vacuoles take up more than 90 percent of the cell volume and grow as the cell grows. The dissolved substances in the vacuole, working together with the vacuolar membrane, provide the turgor or stiffness of the cell which, in turn, provides support for the structure of nonwoody plants. The presence of the dissolved substances causes water to enter the vacuole, making it swell like a balloon. Plant cells have a rigid cell wall which resists the swelling of the vacuole, providing strength in the process.
Reproduction: Some pigments (especially blue and pink ones) in petals and fruits are contained in vacuoles. These pigments—the anthocyanins—are visual cues that help attract the animals that assist in pollination or seed dispersal.
Digestion: In some plants, the vacuoles contain enzymes that hydrolyze seed proteins into monomers that a developing plant embryo can use as food.
Food vacuoles are found in some simple and evolutionarily ancient groups of organisms—single-celled protists and simple multicellular organisms such as sponges, for example. In these organisms, the cells engulf food particles by phagocytosis, generating a food vacuole. Fusion of this vacuole with a lysosome results in digestion and small molecules leave the vacuole and enter the cytoplasm for use or distribution to other organelles.
Contractile vacuoles are found in many freshwater protists. Their function is to get rid of the excess water that rushes into the cell because of the imbalance in salt concentration between the relatively salty interior of the cell and its freshwater environment. The contractile vacuole enlarges as water enters, then abruptly contracts, forcing the water out of the cell through a special pore structure.
Cytoskeleton
In addition to its many membrane-enclosed organelles, the eukaryotic cytoplasm contains a set of long, thin fibers called the cytoskeleton. The cytoskeleton fills at least three important roles:
It maintains cell shape and support.
It provides for various types of cellular movement.
Some of its fibers act as tracks or supports for motor proteins which help move things within the cell.
Microfilaments function in support and movement
Microfilaments can exist as single filaments in bundles or in networks. They are about 7 nm in diameter and several micrometers long. They are assembled from actin, a protein that exists in several forms and has many functions among members of the animal phyla. The actin found in microfilaments (which are also known as actin filaments) is extensively folded and has distinct “head” and “tail” sites. These sites interact with other actin molecules to form long, double helical chains. The polymerization of actin into microfilaments is reversible and they can disappear from cells, breaking down into units of free actin. Microfilaments have two major roles: a)they help the entire cell or parts of the cell to move; b)they stabilize cell shape.
In muscle cells, actin fibers are associated with another protein, myosin, and the interactions of these two proteins account for the contraction of muscles. In non-muscle cells, actin fibers are associated with localized changes of shape in the cell. For example, microfilaments are involved in a flowing movement of the cytoplasm called cytoplasmic streaming and in the “pinching” contractions that divide an animal cell into two daughter cells. Microfilaments are also involved in the formation of cellular extensions called pseudopodia (pseudo-, “false;” podia, “feet”) that enable some cells to move. In some cell types, microfilaments form a meshwork just inside the plasma membrane. Actin-binding proteins then cross-link the microtubules to form a rigid structure that supports the cell. Microfilaments support the tiny microvilli that line the intestine, giving it a larger surface area through which to absorb nutrients.
Intermediate filaments are tough supporting elements
Intermediate filaments are found only in multicellular organisms. In contrast to the other components of the cytoskeleton, there are at least 50 different kinds of intermediate filaments, often specific to a few cell types. They generally fall into six molecular classes, based on amino acid sequence, and share the same general structure being composed of fibrous proteins of the keratin family, similar to the protein that makes up hair and fingernails. In cells, these proteins are organized into tough, ropelike assemblages 8 to 12 nm in diameter. Intermediate filaments have two major structural functions:
They stabilize cell structure.
They resist tension.
In some cells, intermediate filaments radiate from the nuclear envelope and may maintain the positions of the nucleus and other organelles in the cell. The lamins of the nuclear lamina are intermediate filaments. Other kinds of intermediate filaments help hold a complex apparatus of microfilaments in place in muscle cells. Still other kinds stabilize and help maintain rigidity in surface tissues by connecting “spot welds” called desmosomes between adjacent cells.
Microtubules are long and hollow
Microtubules are long, hollow, unbranched cylinders about 25 nm in diameter and up to several micrometers long. Microtubules have two roles in the cell:
They form a rigid internal skeleton for some cells.
They act as a framework along which motor proteins can move structures in the cell.
Microtubules are assembled from molecules of the protein tubulin. Tubulin is a dimer—a molecule made up of two monomers. The polypeptide monomers that make up this protein are known as a-tubulin and b-tubulin. Thirteen chains of tubulin dimers surround the central cavity of the microtubule. The two ends of a microtubule are different. One end is designated the plus (+) end, the other the minus (–) end. Tubulin dimers can be added or subtracted, mainly at the plus end, lengthening or shortening the microtubule. This capacity to change length rapidly makes microtubules dynamic structures. This dynamic property of microtubules is seen in animal cells, where they are often found in parts of the cell that are changing shape. Many microtubules radiate from a region of the cell called the microtubule organizing center. Tubule polymerization results in rigidity and tubule depolymerization leads to a collapse of this rigid structure. In plants, microtubules help control the arrangement of the cellulose fibers of the cell wall. Electron micrographs of plants frequently show microtubules lying just inside the plasma membrane of cells that are forming or extending their cell walls. Experimental alteration of the orientation of these microtubules leads to a similar change in the cell wall and a new shape for the cell. In many cells, microtubules serve as tracks for motor proteins, specialized molecules that use energy to change their shape and move. Motor proteins bond to and move along the microtubules, carrying materials from one part of the cell to another. Microtubules are also essential in distributing chromosomes to daughter cells during cell division. And they are intimately associated with movable cell appendages: the flagella and cilia.
Microtubules power cilia and flagella
Many eukaryotic cells possess flagella or cilia or both. These whiplike organelles may push or pull the cell through its aqueous environment, or they may move surrounding liquid over the surface of the cell. Cilia and eukaryotic (but not prokaryotic) flagella are both assembled from specialized microtubules and have identical internal structures but they differ in their relative lengths and their patterns of beating:
Flagella are longer than cilia and are usually found singly or in pairs. Waves of bending propagate from one end of a flagellum to the other in snakelike undulation.
Cilia are shorter than flagella and are usually present in great numbers. They beat stiffly in one direction and recover flexibly in the other direction (like a swimmer’s arm), so that the recovery stroke does not undo the work of the power stroke.
In cross section, a typical cilium or eukaryotic flagellum is surrounded by the plasma membrane and contains a “9 + 2” array of microtubules. Nine fused pairs of microtubules—called doublets—form an outer cylinder and one pair of unfused microtubules runs up the center. A spoke radiates from one microtubule of each doublet and connects the doublet to the center of the structure. In the cytoplasm at the base of every eukaryotic flagellum or cilium is an organelle called a basal body. The nine microtubule doublets extend into the basal body. In the basal body, each doublet is accompanied by another microtubule, making nine sets of three microtubules. The central, unfused microtubules do not extend into the basal body.
Centrioles are almost identical to the basal bodies of cilia and flagella. Centrioles are found in all eukaryotes except the flowering plants, pine trees and their relatives and some protists. It is made up of a precise bundle of microtubules arranged as nine sets of three fused microtubules each. Centrioles lie in the microtubule organizing center in cells that are about to divide.
Motor proteins move along microtubules
The nine microtubule doublets of cilia and flagella are linkedby proteins. The motion of cilia and flagella results from the sliding of the microtubules past each other. This sliding is driven by a motor protein called dynein, which can undergo changes in its shape. All motor proteins work by undergoing reversible shape changes powered by energy from ATP. Dynein molecules attached to one microtubule doublet bind to a neighboring doublet. As the dynein molecules change shape, they move the microtubule past its neighbor. Dynein and another motor protein, kinesin, are responsible for carrying protein-laden vesicles from one part of the cell to another. These motor proteins bind to a vesicle or other organelle, then “walk” it along a microtubule by changing their shape. Recall that microtubules have a plus end and a minus end. Dynein moves attached organelles toward the minus end, while kinesin moves them toward the plus end.
Extracellular Structures
Although the plasma membrane is the functional barrier between the inside and the outside of a cell, many structures are produced by cells and secreted to the outside of the plasma membrane, where they play essential roles in protecting, supporting or attaching cells. Because they are outside the plasma membrane, these structures are said to be extracellular. The peptidoglycan cell wall of bacteria is such an extracellular structure. In eukaryotes, other extracellular structures—the cell walls of plants and the extracellular matrices found between the cells of multicellular animals—play similar roles. Both of these structures are made up of a prominent fibrous macromolecule embedded in a jellylike medium.
The plant cell wall consists largely of cellulose
The cell wall of plant cells is a semirigid structure outside the plasma membrane (Figure). It consists of cellulose fibers embedded in other complex polysaccharides and proteins. The cell wall has three major roles in plants:
It provides support for the cell and limits its volume by remaining rigid.
It acts as a barrier to infections by fungi and other organisms that can cause plant diseases.
It contributes to plant form by growing as plant cells expand.
Because of their thick cell walls, plant cells viewed under a light microscope appear to be entirely isolated from each other. But electron microscopy reveals that this is not the case. The cytoplasm of adjacent plant cells is connected by numerous plasma membrane-lined channels, called plasmodesmata, that are about 20 to 40 nm in diameter and extend through the walls of adjoining cells. Plasmodesmata permit the diffusion of water, ions, small molecules, and RNA and proteins between connected cells. Such diffusion ensures that the cells of a plant have uniform concentrations of these substances.
Animal cells have elaborate extracellular matrices
The cells of multicellular animals lack the semirigid cell wall that is characteristic of plant cells, but many animal cells are surrounded by, or are in contact with, an extracellular matrix. This matrix is composed of fibrous proteins such as collagen (the most abundant protein in mammals) and glycoproteins. These proteins, along with other substances that are specific to certain body tissues, are secreted by cells that are present in or near the matrix. The functions of the extracellular matrix are many:
It holds cells together in tissues.
It contributes to the physical properties of cartilage, skin, and other tissues.
It helps filter materials passing between different tissues.
It helps orient cell movements during embryonic development and during tissue repair.
It plays a role in chemical signaling from one cell to another.
In the human body, some tissues, such as those in the brain, have very little extracellular matrix; other tissues, such as bone and cartilage, have large amounts. Bone cells are embedded in an extracellular matrix that consists primarily of collagen and calcium phosphate. This matrix gives bone its familiar rigidity. Epithelial cells, which line body cavities, lie together as a sheet spread over a basal lamina, or basement membrane, a form of extracellular matrix.
Some extracellular matrices are made up, in part, of an enormous proteoglycan. A single molecule of this proteoglycan consists of many hundreds of polysaccharides covalently attached to about a hundred proteins, all of which are attached to one enormous polysaccharide. The molecular weight of this proteoglycan can exceed 100 million; the molecule takes up as much space as an entire prokaryotic cell.
How Things Get Into & Out Of Cells
Cells are able to regulate the passage of materials across cell membranes. This is an important capacity. One of the criteria by which we identify living systems is that living matter, although surrounded on all sides by nonliving matter, is different from it in the kinds & amounts of chemical substances it contains. Without this difference, of course, living matter would be unable to maintain the organization & structure on which its existence depends. The cell membrane is not simply an impenetrable barrier, however. Living matter constantly exchanges substances with the nonliving world around it. Control of these exchanges is essential in order to protect the cell’s integrity & to maintain those very narrow conditions of pH and salt concentrations, common to all cells, at which enzyme activity can take place. The cell membrane, thus, has a complex double function of keeping things out & letting things in. Moreover, cell membranes not only control the passage of material from outside the cell, but internal membranes, such as those surrounding mitochondria, chloroplast and the nucleus, regulate the passage of materials between intracellular compartment & so regulate their internal environment.
The regulation of substances moving across membranes depends on interactions between the physical & chemical properties of the membrane & those of the molecules that penetrate through them. Of the many kinds of molecules moving in & out of cells, the most important is water. Let us therefore look again at water, focusing our attention this time on how water moves.
Moving processes (passive transport)
Three principles govern the movement of substances: bulk flow, diffusion, and osmosis.
Bulk flow is the overall movement of water (or some other liquid). It occurs in response to differences in the potential energy of water, usually referred to as water potential.
A simple example of water that has potential energy is water at the top of a hill. As this water runs downhill, its potential energy can be converted to mechanical energy by a watermill or to electrical energy by a hydroelectric turbine.
Water moves from an area where water potential is greater to an area where water potential is less regardless of the reason for the water potential. Your heart pumps blood to your brain against gravity by creating a greater water (blood) potential.
Molecules constantly move. They constantly bumper into and bounce off one another. Life scientists call the movement of molecules from a crowded area to less crowded area diffusion. In other words diffusion is the movement of suspended or dissolved particles from a more concentrated to a less concentrated region as a result of the random movement of individual particles; the process tends to distribute them uniformly throughout a medium.
When drops of red ink are added to a glass of water, the ink molecules are very close together. Diffusion takes place as the ink molecules move through the water. In time, the ink molecules spread out evenly until all of the water is of a light red color.
A permeable membrane of the cell has small holes through which molecules can move. Diffusion will take place if ink molecules are more crowded on one side of a permeable membrane, than on the other. The ink molecules will diffuse through the permeable membrane until they are evenly spaced on both sides of the membrane.
The holes in a cell membrane allow only some molecules to diffuse through, but not all. For this reason, life scientists describe a cell membrane as semipermeable. Water and other small molecules can easily pass through a cell membrane. Proteins and other large molecules must be broken down into smaller parts to get through a cell membrane. Once inside a cell, the larger molecules can be rebuilt.
Every cell continually uses sugar and blood must bring new supplies of sugar to cells. Sugar molecules diffuse in through the cell membrane, because there are fewer sugar molecules on the inside of the cell than on the outside. The cells also continually produce wastes. These wastes diffuse out of the cell through the semipermeable cell membrane because there are fewer waste molecules outside the cell than inside (Fig. 2.1). As the blood carries the wastes away, more wastes can diffuse out of the cells.
The diffusion of water into and out of the cell is known as osmosis. The process can cause a cell to swell or to shrink depending on the amount of water around the cell. The effects of osmosis can be easily seen in large cells, such as yolks of chicken eggs.
Osmosis can cause shrinking and swelling in plants too. When plant cells swell because of extra water in their environment, they push against their cell walls. This keeps the stems and leaves upright. When plant cells lack water in their environment, they shrink away from their cell walls, this makes a plant wilt.
Pumping Molecules Through Cell Membranes (active transport)
Sometimes a cell uses energy to pump molecules through its cell membrane. This process - active transport (Fig. 2.2) - can move molecules and ions from less crowded area to more crowded areas. In other words, active transport can move molecules away from the direction in which they diffuse.
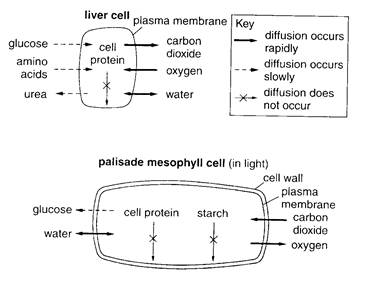
Fig. 2.1. Diffusion into and out of cells
The cell membrane probably contains special molecules that act as carriers for active transport. The carrier molecules can pick up the molecules from outside the cell membrane, pump them through the membrane, and release them on the inside of the cell. This energy process might be similar to that of a ferryboat bringing people across a body of water.
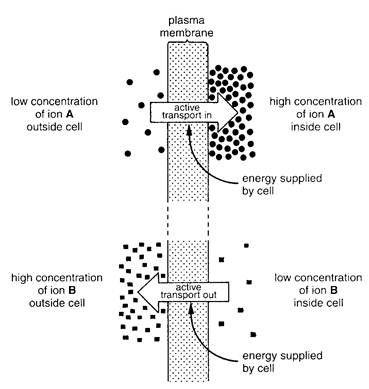
Fig. 2.2. Active transport of two different ions
Date: 2014-12-22; view: 1561
| <== previous page | | | next page ==> |
| ELEMENTS OF GENERAL BIOLOGY | | | Energy, Enzymes, and Metabolism |