CATEGORIES:
BiologyChemistryConstructionCultureEcologyEconomyElectronicsFinanceGeographyHistoryInformaticsLawMathematicsMechanicsMedicineOtherPedagogyPhilosophyPhysicsPolicyPsychologySociologySportTourism
Metapopulations and migration
Populations are also studied and modeled according to the metapopulation concept. The metapopulation concept was introduced in 1969:[56] "as a population of populations which go extinct locally and recolonize."[57]:105 Metapopulation ecology is another statistical approach that is often used in conservation research.[58] Metapopulation research simplifies the landscape into patches of varying levels of quality.[59] Metapopulations are linked by the migratory behaviours of organisms. Animal migration is set apart from other kinds of movement because it involves the seasonal departure and return of individuals from one habitat to another.[60] Migration is also a population level phenomenon, such as the migration routes followed by plants as they occupied northern post-glacial environments. Plant ecologists rely on pollen records that accumulate and stratify in wetlands to reconstruct the timing of plant migration and dispersal relative to historic and contemporary climates. These migration routes involved an expansion of the range as plant populations expanded from one area to another. There is a larger taxonomy of movement, such as commuting, foraging, territorial behaviour, stasis, and ranging. Dispersal is usually distinguished from migration because it involves the one way permanent movement of individuals from their birth population into another population.[61][62]
In metapopulation terminology there are emigrants (individuals that leave a patch), immigrants (individuals that move into a patch) and sites are classed either as sources or sinks. A site is a generic term that refers to places where ecologists sample populations, such as ponds or defined sampling areas in a forest. Source patches are productive sites that generate a seasonal supply of juveniles that migrate to other patch locations. Sink patches are unproductive sites that only receive migrants and will go extinct unless rescued by an adjacent source patch or environmental conditions become more favorable. Metapopulation models examine patch dynamics over time to answer questions about spatial and demographic ecology. The ecology of metapopulations is a dynamic process of extinction and colonization. Small patches of lower quality (i.e., sinks) are maintained or rescued by a seasonal influx of new immigrants. A dynamic metapopulation structure evolves from year to year, where some patches are sinks in dry years and become sources when conditions are more favorable. Ecologists use a mixture of computer models and field studies to explain metapopulation structure.[63][64]
Community ecology

Interspecific interactions such as predation are a key aspect of community ecology.
Community ecology examines how interactions among species and their environment affect the abundance, distribution and diversity of species within communities.
Community ecology is the study of the interactions among a collection of interdependent species that cohabitate the same geographic area. An example of a study in community ecology might measure primary production in a wetland in relation to decomposition and consumption rates. This requires an understanding of the community connections between plants (i.e., primary producers) and the decomposers (e.g., fungi and bacteria).[66] or the analysis of predator-prey dynamics affecting amphibian biomass.[67] Food webs and trophic levels are two widely employed conceptual models used to explain the linkages among species.[68][69]
Ecosystem ecology
These ecosystems, as we may call them, are of the most various kinds and sizes. They form one category of the multitudinous physical systems of the universe, which range from the universe as a whole down to the atom.


Figure 1. A riparian forest in the White Mountains, New Hampshire (USA), an example of ecosystem ecology
The concept of the ecosystem was fully synthesized in 1935 to describe habitats within biomes that form an integrated whole and a dynamically responsive system having both physical and biological complexes. However, the underlying concept can be traced back to 1864 in the published work of George Perkins Marsh ("Man and Nature").[71][72] Within an ecosystem there are inseparable ties that link organisms to the physical and biological components of their environment to which they are adapted.[70] Ecosystems are complex adaptive systems where the interaction of life processes form self-organizing patterns across different scales of time and space.[73] terrestrial, freshwater, atmospheric, and marine ecosystems very broadly cover the major types. Differences stem from the nature of the unique physical environments that shapes the biodiversity within each. A more recent addition to ecosystem ecology are the novel technoecosystems of the anthropocene.[6]
Food webs
A food web is the archetypal ecological network. Plants capture and convert solar energy into the biomolecular bonds of simple sugars during photosynthesis. This food energy is transferred through a series of organisms starting with those that feed on plants and are themselves consumed. The simplified linear feeding pathways that move from a basal trophic species to a top consumer is called the food chain. The larger interlocking pattern of food chains in an ecological community creates a complex food web. Food webs are a type of concept map or a heuristic device that is used illustrate and study pathways of energy and material flows.[7][74][75]
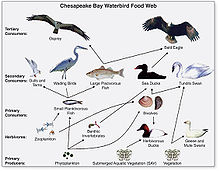
Generalized food web of waterbirds from Chesapeake Bay
Food webs are often limited relative to the real world. Complete empirical measurements are generally restricted to a specific habitat, such as a cave or a pond. Principles gleaned from food web microcosm studies are used to extrapolate smaller dynamic concepts to larger systems.[76] Feeding relations require extensive investigations into the gut contents of organisms, which can be very difficult to decipher, or (more recently) stable isotopes can be used to trace the flow of nutrient diets and energy through a food web.[77] While food webs often give an incomplete measure of ecosystems, they are nonetheless a valuable tool in understanding community ecosystems.[78]
Food-webs exhibit principals of ecological emergence through the nature of trophic entanglement, where some species have many weak feeding links (e.g., omnivores) while some are more specialized with fewer stronger feeding links (e.g., primary predators). Theoretical and empirical studies identify non-random emergent patterns of few strong and many weak linkages that serve to explain how ecological communities remain stable over time.[79] Food-webs have compartments, where the many strong interactions create subgroups among some members in a community and the few weak interactions occur between these subgroups. These compartments increase the stability of food-webs.[80] As plants grow, they accumulate carbohydrates and are eaten by grazing herbivores. Step by step lines or relations are drawn until a web of life is illustrated.
Trophic levels
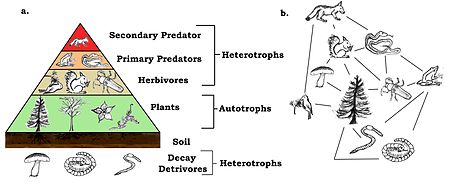
A trophic pyramid (a) and a food-web (b) illustrating ecological relationships among creatures that are typical of a northern Boreal terrestrial ecosystem. The trophic pyramid roughly represents the biomass (usually measured as total dry-weight) at each level. Plants generally have the greatest biomass. Names of trophic categories are shown to the right of the pyramid. Some ecosystems, such as many wetlands, do not organize as a strict pyramid, because aquatic plants are not as productive as long-lived terrestrial plants such as trees. Ecological trophic pyramids are typically one of three kinds: 1) pyramid of numbers, 2) pyramid of biomass, or 3) pyramid of energy.[6]
The Greek root of the word troph, τροφή, trophē, means food or feeding. Links in food-webs primarily connect feeding relations or trophism among species. Biodiversity within ecosystems can be organized into vertical and horizontal dimensions. The vertical dimension represents feeding relations that become further removed from the base of the food chain up toward top predators. A trophic level is defined as "a group of organisms acquiring a considerable majority of its energy from the adjacent level nearer the abiotic source."[84]:383 The horizontal dimension represents the abundance or biomass at each level.[85] When the relative abundance or biomass of each functional feeding group is stacked into their respective trophic levels they naturally sort into a 'pyramid of numbers'.[86]
Functional groups are broadly categorized as autotrophs (or primary producers), heterotrophs (or consumers), and detrivores (or decomposers). Autotrophs are organisms that can produce their own food (production is greater than respiration) and are usually plants or cyanobacteria that are capable of photosynthesis but can also be other organisms such as bacteria near ocean vents that are capable of chemosynthesis. Heterotrophs are organisms that must feed on others for nourishment and energy (respiration exceeds production).[6] Heterotrophs can be further sub-divided into different functional groups, including: primary consumers (strict herbivores), secondary consumers (carnivorous predators that feed exclusively on herbivores) and tertiary consumers (predators that feed on a mix of herbivores and predators).[87] Omnivores do not fit neatly into a functional category because they eat both plant and animal tissues. It has been suggested that omnivores have a greater functional influence as predators because relative to herbivores they are comparatively inefficient at grazing.[88]
Trophic levels are part of the holistic or complex systems view of ecosystems.[89][90] Each trophic level contains unrelated species that grouped together because they share common ecological functions. Grouping functionally similar species into a trophic system gives a macroscopic image of the larger functional design.[91] While the notion of trophic levels provides insight into energy flow and top-down control within food webs, it is troubled by the prevalence of omnivory in real ecosystems. This has lead some ecologists to "reiterate that the notion that species clearly aggregate into discrete, homogeneous trophic levels is fiction."[92]:815 Nonetheless, recent studies have shown that real trophic levels do exist, but "above the herbivore trophic level, food webs are better characterized as a tangled web of omnivores."[93]:612
Keystone species
A keystone species is a species that is disproportionately connected to more species in the food-web. Keystone species have lower levels of biomass in the trophic pyramid relative to the importance of their role. The many connections that a keystone species holds means that it maintains the organization and structure of entire communities. The loss of a keystone species results in a range of dramatic cascading effects that alters trophic dynamics, other food-web connections and can cause the extinction of other species in the community.[94][95]
Sea otters (Enhydra lutris) are commonly cited as an example of a keystone species because they limit the density of sea urchins that feed on kelp. If sea otters are removed from the system, the urchins graze until the kelp beds disappear and this has a dramatic effect on community structure.[96] Hunting of sea otters, for example, is thought to have indirectly led to the extinction of the Steller's Sea Cow (Hydrodamalis gigas).[97] While the keystone species concept has been used extensively as a conservation tool, it has been criticized for being poorly defined from an operational stance. It is very difficult to experimentally determine in each different ecosystem what species may hold a keystone role. Furthermore, food-web theory suggests that keystone species may not be all that common. It is therefore unclear how generally the keystone species model can be applied.[96][98]
Soils
Soil is the living top layer of mineral and organic dirt that covers the surface of the planet, it is the chief organizing centre of most ecosystem functions, and it is of critical importance in agricultural science and ecology. The decomposition of dead organic matter, such as leaves falling on the forest floor, turns into soils containing minerals and nutrients that feed into plant production. The total sum of the planet's soil ecosystems is called the pedosphere where a very large proportion of the Earth's biodiversity sorts into other trophic levels. Invertebrates that feed and shred larger leaves, for example, create smaller bits for smaller organisms in the feeding chain. Collectively, these are the detrivores that regulate soil formation. [99][100][101][102] Tree roots, fungi, bacteria, worms, ants, beetles, centipedes, spiders, mammals, birds, reptiles, amphibians and other less familiar creatures all work to create the trophic web of life in soil ecosystems. As organisms feed and migrate through soils they physically displace materials, which is an important ecological process called bioturbation. Bioturbation helps to aerate the soils, thus stimulating hetertrophic growth and production. Biomass of soil microorganisms are influenced by and feed back into the trophic dynamics of the exposed solar surface ecology. Paleoecological studies of soils places the origin for bioturbation to a time before the Cambrian period. Other events, such as the evolution of trees and amphibians moving into land in the Devonian period played a significant role in the development of the ecological trophism in soils.[67][102][103]
Date: 2016-01-03; view: 4053
| <== previous page | | | next page ==> |
| Hierarchical ecology | | | Ecological complexity |