CATEGORIES:
BiologyChemistryConstructionCultureEcologyEconomyElectronicsFinanceGeographyHistoryInformaticsLawMathematicsMechanicsMedicineOtherPedagogyPhilosophyPhysicsPolicyPsychologySociologySportTourism
Fig. 22.11. Reproduction
Sexual Reproduction
Sexual reproduction in many club fungi mushrooms begins in the same way as it does for members of the two fungal phyla previously discussed. When a spore lands in a suitable place - often an area with good organic material and humus in the soil - it germinates and produces a mycelium just beneath the surface. The hyphae of the mycelium are divided into cells that each contain a single haploid nucleus. Such a mycelium is said to be monokaryotic. Monokaryotic mycelia of club fungi often occur in four mating types, usually designated simply as types 1, 2, 3, and 4. Only types 1 and 3, or types 2 and 4, can mate with each other. If the growth of the hyphae of compatible mating types happens to bring them close together, cells of each mycelium may unite, initiating a new mycelium in which each cell has two nuclei. Such a mycelium is said to be dikaryotic. Dikaryotic mycelia sometimes have little walled-off bypass loops called clamp connections between cells on the surface of the hyphae. The clamp connections develop as a result of a unique type of mitosis that ensures each cell will have one nucleus of each original mating type within it.
After developing for a while, the dikaryotic mycelium may become very dense and form a compact, solid-looking mass called a button. This pushes above the surface and expands into a basidioma (formerly known as a basidiocarp), commonly called a mushroom. Most mushrooms have an expanded umbrellalike cap (pileus) and a stalk (stipe). Some have a ring called an annulus on the stalk. It is the remnant of a membrane that extended from the cap to the stalk and tore as the cap expanded. Some mushrooms, such as the notorious death angel and destroying angel mushrooms (Amanita spp.), also have a cup called a volva at the base. Thin, fleshy-looking plates called gills radiate out from the stalk on the underside of the cap. Microscopic examination of a gill reveals it is composed of compacted hyphae, with large numbers of basidia oriented at right angles to the flat surfaces of the gill.
As each basidium matures, the two nuclei unite, and the diploid nucleus undergoes meiosis. The four nuclei that result from meiosis migrate through four (in a few species, two) tiny pegs at the tip of the basidium, walls forming around the nuclei in the process. The resultant cells are now basidiospores. The tiny pegs, called sterigmata (singular: sterigma), serve as stalks for the basidiospores. One large mushroom may produce several billion basidiospores within a few days. These are forcibly discharged into the air between the gills. They then drift downward and blow away with the slightest breeze. If you remove a mushroom stalk and place the cap gill-side down on a piece of paper, covering it with a dish to eliminate air currents, the spores will fall and adhere to the paper in a pattern perfectly reflecting the arrangement of the gills. The dish and cap can be removed a day later, and the spore print can be made more or less permanent with the application of clear varnish or shellac. Such spore prints can be used as an aid to identification, employing white paper for dark-colored spores and black paper for white or light-colored spores.In nature, some of the basidiospores eventually repeat the reproductive cycle. Often a dikaryotic mycelium radiates out from its starting point, periodically producing basidiomata in so-called fairy rings. If conditions are favorable, the mycelium continues to grow at the edges for many years while dying in the center as food resources are depleted. Some mycelia have been known to grow in this way for over 500 years.
LICHENS
Lichens traditionally have been referred to as prime examples of symbiotic relationships. Each consists of a fungus and an alga (or cyanobacterium) intimately associated in a spongy thallus. The thallus can range in diameter from less than 1 millimeter to more than 2 meters (0.04 inch to 6.5 feet). The photosynthetic component supplies the food for both organisms. The fungus protects the photosynthetic organisms from harmful light intensities, produces a substance that accelerates the rate of photosynthesis, and absorbs and retains water and minerals for both organisms. Supporting the belief that a lichen involves a symbiotic relationship is the fact that neither the fungus nor the photosynthetic organisms grow independently where the lichen grows. The physiological evidence suggests, however, that it would probably be more correct to say that the fungus parasitizes the photosynthetic component in a controlled fashion, actually destroying chlorophyll-containing cells in some instances.
There are about 14,500 known species of lichens. The photosynthetic component is either a green alga or acyanobacterium, a few lichens having two species of algae present. Three genera of green algae and one genus of cyanobacterium are involved in 90% of all lichen species, and one species of alga may be found in many different lichens.
Each lichen, however, has its own unique species of fungus. With the exception of about 20 tropical species of lichens that have a club fungus and one species (associated with bald cypress trees) that has a bacterial component, lichens have members of the sac fungi for their fungal components. It is possible to isolate and culture the components separately. When this is done, however, the fungus takes on a very different, compact but indefinite shape, and the algae or cyanobacteria grow faster than they do when they are part of a lichen. The fungal component is very rarely found growing independently in nature, while the photosynthetic component is known to thrive independently of the lichen in some instances. Lichen species, therefore, are identified according to the fungus present.
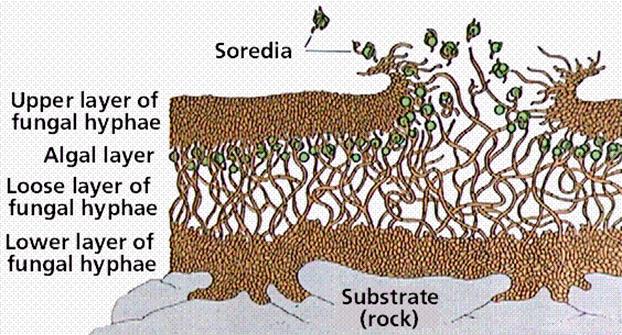
Fig.22.12. Lichens
Lichens grow very slowly, at a maximum rate of 1 centimeter (0.4 inch) and a minimum of as little as 0.1 millimeter (0.004 inch) per year. They are capable of living to an age of 4,500 or more years and are tolerant of environmental conditions that kill most other forms of life. They are found on bare rocks in the blazing sun or bitter cold in deserts, in both arctic and antarctic regions, on trees, and just below the permanent snow line of high mountains where nothing else will grow. One species grows completely submerged on ocean rocks. Lichens even attach themselves to manufactured substances, such as glass, concrete, and asbestos. Part of the reason for the wide range of tolerance of lichens is the presence in the lichens’ thalli of gelatinous substances, which enable them to withstand periods of rapid drying alternating with wet spells. While they are dry, their water content may drop to as low as 2% of their dry weight, and the upper part of the thallus becomes opaque enough to exclude much of the light that falls on them. In this state, most environmental extremes do not affect them at all, as they temporarily become dormant and do not carry on photosynthesis.The lichen thallus usually consists of three or four layers of cells or hyphae. At the surface is a protective layer constituting the upper cortex.

Fig.22.13. Lichens
1. Upper cortex:Under magnification, a section through a typical foliose lichen thallus reveals four layers of interlaced fungal filaments. The uppermost layer is formed by densely agglutinated fungal hyphae building a protective outer layer called the cortex, which can reach several hundred μm in thickness. Beneath the upper cortex is an algal layer composed of algal cells embedded in rather densely interwoven fungal hyphae.
2. Algae layer: This part makes up 5% of the plant. It provides food for the thallus (i.e. photosynthesises) and is light loving.
3. Fungal Hyphae: The fungal part provides moisture and shelter, it is therefore the dominant partner. The thallus consists of 95% fungi.
4. Medulla: Beneath this algal layer is a third layer of loosely interwoven fungal hyphae without algal cells. This layer is called the medulla.
5. Lower cortex: Beneath the medulla, the bottom surface resembles the upper surface and is called the lower cortex, again consisting of densely packed fungal hyphae. The lower cortex often bears rootlike fungal structures known as rhizines, which serve to attach the thallus to the substrate on which it grows
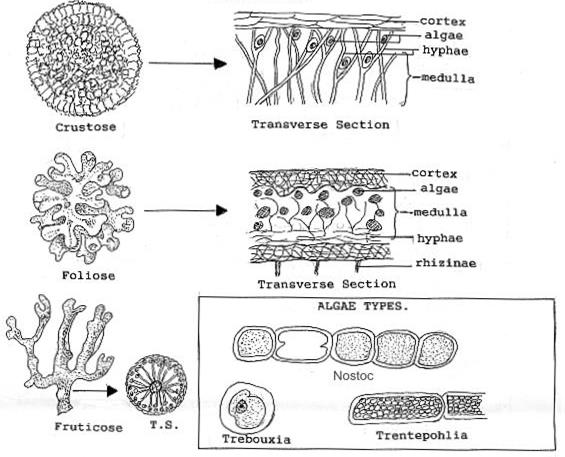
The hyphae are so compressed they resemble parenchyma cells, and it is here that the gelatinous substances aid in the layer’s protective function. Below the upper cortex is an algal layer in which algal (or cyanobacterial) cells are scattered among strands of hyphae. Next is a medulla consisting of loosely packed hyphae, which occupies at least half the volume of the thallus. A number of substances produced by the lichen are stored here. A fourth layer, called the lower cortex, is frequently but not always present. It resembles the upper cortex but is usually thinner and is often covered with anchoring strands of hyphae called rhizines. Lichens have been loosely grouped into three major growth forms, which have no basis in their natural relationships but are convenient as a first step in their identification. Crustose lichens are attached to or embedded in their substrate over their entire lower surface. They often form brightly colored crusty patches on bare rocks and tree bark. The hyphae of some that grow on sedimentary rocks penetrate as much as 1 centimeter (0.4 inch) into the rock. Others grow just beneath the cuticle of the leaves of tropical hardwood trees, with no apparent harm to the leaves.
Date: 2016-01-03; view: 2114
| <== previous page | | | next page ==> |
| Phylum Basidiomycota—The Basidiomycetes (Club Fungi) | | | Fig.22.14. Lichens Type |